1.Kvartal: Den mest interessante biologi-artikkelen fra 2022 (så langt)
Oversatt herfra. (kursiv og understreking ved oversetter)
Bilde 1. Darwins biller
 Vi har nettopp avsluttet årets første kvartal. Det er lenge til nyttårsaften 2022. Men denne nye åpne artikkelen fra seniorforfatter Sara Walker (Arizona State) og hennes samarbeidspartnere vil være vanskelig å toppe, i "Wow, det er så interessant!" kategori. (Den første forfatteren av denne artikkelen er Dylan Gagler, så vi vil referere til den som "Gagler et al. 2022" nedenfor.)
Vi har nettopp avsluttet årets første kvartal. Det er lenge til nyttårsaften 2022. Men denne nye åpne artikkelen fra seniorforfatter Sara Walker (Arizona State) og hennes samarbeidspartnere vil være vanskelig å toppe, i "Wow, det er så interessant!" kategori. (Den første forfatteren av denne artikkelen er Dylan Gagler, så vi vil referere til den som "Gagler et al. 2022" nedenfor.)
1. På den tiden var det beste beviset for et enkelt livstre, forankret i den siste universelle felles stamfaren (LUCA), den tilsynelatende biokjemiske og molekylære universaliteten til jordisk liv.
Den ledende neo-darwinistiske Theodosius Dobzhansky uttrykte dette poenget veltalende i sitt berømte essay fra 1973, "Ingenting i biologi gir mening bortsett fra i lys av evolusjon":
Livets enhet er ikke mindre bemerkelsesverdig enn dets mangfold...Ikke bare er DNA-RNA genetisk kode universell, men det er også metoden for oversettelse av sekvensene av "bokstavene" i DNA-RNA til sekvenser av aminosyrer i proteiner. De samme 20 aminosyrene utgjør utallige forskjellige proteiner i alle, eller i det minste i de fleste, organismer.  Ulike aminosyrer er kodet av en til seks nukleotidtripletter i DNA og RNA. Og de biokjemiske universalene strekker seg utover den genetiske koden og dens oversettelse til proteiner: slående uniformiteter råder i celle-metabolismen til de mest forskjellige levende vesener. Adenosintrifosfat, biotin, riboflavin, hemes, pyridoksin, vitamin K og B12, og folsyre implementerer metabolske prosesser overalt. Hva betyr disse biokjemiske eller biologiske universellene? De antyder at liv oppsto fra livløs materie bare én gang, og at alle organismer, uansett hva de nå er forskjellige, i andre henseender, bevarer de grunnleggende trekk ved urlivet.[Uthevelse tilføyd.]
Ulike aminosyrer er kodet av en til seks nukleotidtripletter i DNA og RNA. Og de biokjemiske universalene strekker seg utover den genetiske koden og dens oversettelse til proteiner: slående uniformiteter råder i celle-metabolismen til de mest forskjellige levende vesener. Adenosintrifosfat, biotin, riboflavin, hemes, pyridoksin, vitamin K og B12, og folsyre implementerer metabolske prosesser overalt. Hva betyr disse biokjemiske eller biologiske universellene? De antyder at liv oppsto fra livløs materie bare én gang, og at alle organismer, uansett hva de nå er forskjellige, i andre henseender, bevarer de grunnleggende trekk ved urlivet.[Uthevelse tilføyd.]
For Dobzhansky, som for alle ny-darwinister (per definisjon), bekreftet den tilsynelatende molekylære universaliteten til livet på jorden Darwins spådom om at alle organismer "har nedstammet fra en urform, som liv først ble pustet inn i" (1859, 494) - en enhet som nå er kjent som Last Universal Common Ancestor, eller LUCA. Så sterk kraften av denne tilsynelatende universaliteten, forankret i LUCA, at enhver annen historisk geometri virker utenkelig.
Bilde 2. Gruppering av metabolske prosesser
"Livets lover"
Teoretiker Sara Walker og hennes team av samarbeidspartnere leter imidlertid etter en redegjørelse for det de kaller (i Gagler et al. 2022) "livets lover" som vil gjelde "for alle mulige biokjemier" - inkludert organismer som finnes andre steder i universet, hvis det eksisterer. For det formål ønsket de å vite om den molekylære universaliteten forklart under neo-darwinistisk teori som materiell avstamning fra LUCA (a) virkelig eksisterer, og (b) hvis ikke, hvilke mønstre eksisterer, og hvordan kan de forklares uten å forutsette en enkelt felles stamfar.
Og en enkelt felles stamfar, LUCA? Det var det de ikke fant.
2. Tell opp de forskjellige enzymfunksjonene - og kartlegg deretter det tallet innenfor det totale funksjonelle rommet.
Mange tusen forskjellige funksjonelle enzymklasser, nødvendige for den levende tilstanden, er blitt beskrevet og katalogisert i Enzyme Commission Classification, i henhold til deres utpekte EC-nummer. Disse betegnelsene har fire sifre, som tilsvarer gradvis mer spesifikke funksjonsklasser. Tenk for eksempel på enzymet tyrosin-tRNA-ligase. Dets EC-nummer, 6.1.1.1, indikerer et nestet sett med klasser: EC 6 omfatter ligasene (bindingsdannende enzymer); EC 6.1, de ligasene som danner karbon-oksygenbindinger; 6.1.1, ligaser som danner aminoacyl-tRNA og beslektede forbindelser; til slutt, 6.1.1.1, de spesifikke ligasene som danner tyrosin-tRNA. (Se figur 1.)
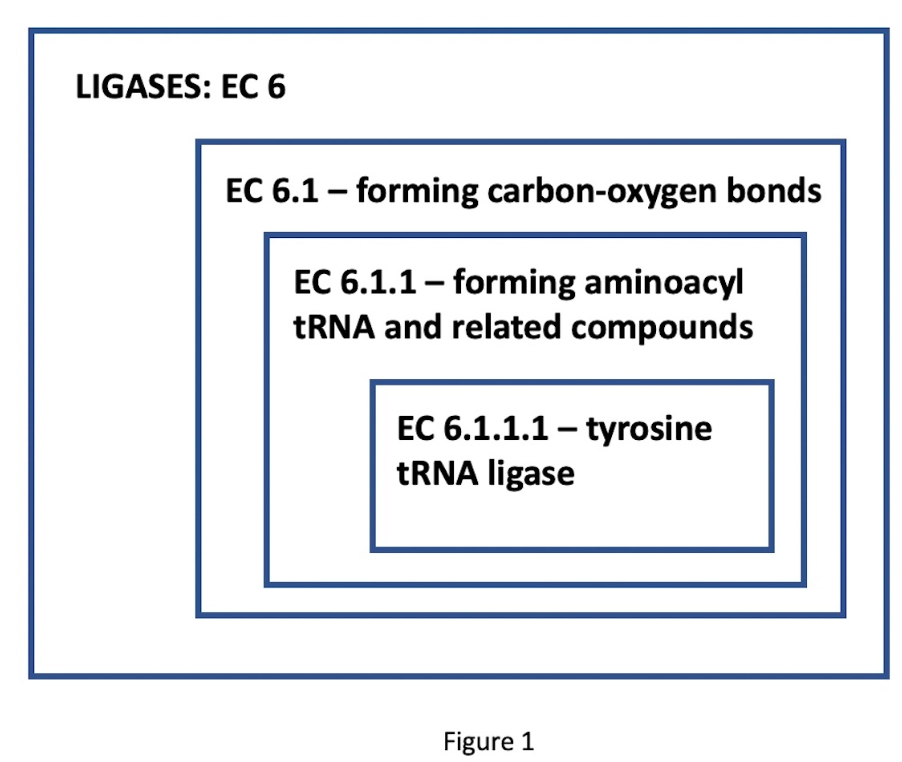 Bilde 3. Figur 1
Bilde 3. Figur 1
Det viktigste takeaway fra dette mønsteret?
å være en ligase - nemlig et enzym som danner bindinger ved hjelp av ATP - innebærer å tilhøre en funksjonell gruppe, men ikke en gruppe med materiell identitet blant medlemmene. En grov parallell til et naturlig språk som norsk kan være nyttig. Anta at du ønsket å uttrykke ideen om "mørke" eller "mørket" (dvs. det relative fraværet av lys). Norsk gir et bredt spekter av synonymer for "mørket", for eksempel: grumsete; skyggelagt; nedtonet; tilslørt
Det samme ville være tilfelle - eksistensen av et sett med synonymer, dvs. ord med samme generelle betydning, men ikke samme sekvensidentitet - for enhver annen idé. Konseptet med at noe blir "blokkert", for eksempel, har synonymene:
fastkjørt; fastlåst; okkludert; forhindret; hindret
Selv om disse ordene formidler (omtrent) samme betydning, og dermed faller inn i de samme semantiske funksjonelle klassene, er de ikke de samme tegnstrengene. Plasseringene deres i en ordbok, sortert etter alfabetsekvens, kan være hundrevis av sider fra hverandre. Videre, som studert av disiplinen komparativ filologi, vil de historiske røttene til et ord som "hindret" avvike radikalt fra dets funksjonelle synonymer, som "blokkert". Disse to ordene, selv om de semantisk i stor grad er synonyme, kommer inn på norsk fra opprinnelig divergerende eller urelaterte forløpere - et tegnstrenggap som fortsatt gjenspeiles av deres svært forskjellige stavemåter.
Et slående likt mønster oppnås med de kritiske (essensielle) komponentene til alle organismer. Gagler et al. 2022 så på overflod av enzymfunksjoner på tvers av de tre hoveddomenene i livet (bakterier, Archaea, Eukarya), så vel som i metagenomer (miljøprøvet DNA). Det de fant var bemerkelsesverdig - et funn (se nedenfor) som kan være lettere for ikke-biologiske lesere å forstå via en annen analogi.
3. En glidende overgang i dataarkitekturer - så tilbake til enzymer.
Den grunnleggende arkitekturen til bærbare datamaskiner inkluderer komponenter som finnes i en slik maskin, definert av deres funksjonelle roller:
Central Processing Unit (CPU) - Sentral prosessor enheten
Minne - lagring av kodet informasjon
Strømforsyning - elektroner (energi) som trengs for at noe i det hele tatt skal kunne beregnes
Og så videre. (Selv om å utforske dette punktet i detalj ville ta oss langt til siden, er det verdt å merke seg at i 1936, da Alan Turing definerte en universell beregningsmaskin, gjorde han det uten noen anelse om ankomsten, flere tiår i forkant, av silisiumbasert integrert kretser, miniatyriserte transistorer, hovedkort, solid-state minneenheter eller noen av de andre materielle delene av datamaskiner som vi nå er så kjent med. Snarere var delene hans funksjonelle, ikke-materielt definert, som abstraksjoner som okkuperte de forskjellige rollene disse delene ville spille i beregningsprosessen - uansett hvilken materiell instansiasjon deres senere skulle vise seg å være.) Anta nå at vi undersøkte 100 000 bærbare datamaskiner, tilfeldig samplet fra hele USA, for å se hvilken type CPU - som betyr hvilken materialdel (f.eks. bygget av hvilken produsent) - hver enkelt maskin brukt som primær logikkoperatør.
 En rekke utfall er mulig (se figur 2A og 2B). For eksempel, hvis vi plotter CPU-er fra forskjellige produsenter på y-aksen, mot det totale antallet bærbare deler inspisert på x-aksen, kan det være tilfelle at fordelingen av forskjellig produserte (dvs. materielt distinkte) CPUer vil skaleres lineært med bærbare datamaskiner inspisert (Figur 2A). Med andre ord, etter hvert som vårt utvalg av inspiserte bærbare deler vokser, vil antallet forskjellige CPUer som oppdages, helle tilsvarende oppover.
En rekke utfall er mulig (se figur 2A og 2B). For eksempel, hvis vi plotter CPU-er fra forskjellige produsenter på y-aksen, mot det totale antallet bærbare deler inspisert på x-aksen, kan det være tilfelle at fordelingen av forskjellig produserte (dvs. materielt distinkte) CPUer vil skaleres lineært med bærbare datamaskiner inspisert (Figur 2A). Med andre ord, etter hvert som vårt utvalg av inspiserte bærbare deler vokser, vil antallet forskjellige CPUer som oppdages, helle tilsvarende oppover.
Eller - og dette passer selvfølgelig med den faktiske situasjonen vi finner (se figur 2B) - de fleste bærbare datamaskiner vil inneholde prosessorer produsert enten av Intel eller AMD. I dette tilfellet ville vi plotte en linje hvis helning ville endre seg mye saktere, og holde seg stort sett flat, faktisk etter at CPU-ene fra Intel og AMD ble talt opp.
Bilde 4. Enzym-dannelse
Kjerne-begrunnelsen for deres tilnærming
Vurder nå figur 3 (nedenfor), fra Gagler et al. 2022 artikkelen -beskrevet her. Dette viser kjerne-begrunnelsen for tilnærmingen deres: tell de EC-klassifiserte enzym 'delene' innenfor hvert av hoveddomenene, og fra metagenomer, og plott deretter dette sammen mot de totale EC-tallene.
Figur 3 viser også hovedfunnet deres. Ettersom enzym-reaksjonsrommet vokser (på den horisontale aksen - totale EC-tall), vokser også antallet unike funksjoner (på den vertikale aksen - EC-antall i hver EC-klasse).
Lærdommen som Gagler et al. 2022 trekke fra denne oppdagelsen? Mønsteret skyldes IKKE materiale som stammer fra en enkelt felles stamfar, LUCA. Faktisk, under overskriften "Universalitet i skalering av enzymfunksjon er ikke forklart av universelt delte komponenter," forklarer de at materiell avstamning fra LUCA vil innebære delte "mikroskalatrekk", som betyr "spesifikke molekyler og reaksjoner som brukes av alt liv," eller "delt komponentkjemi på tvers av systemer." Hvis vi bruker CPU/laptop-analogien, vil denne mikroskala-fellesskapet tilsvare å finne CPUer fra samme produsent, med de samme interne logiske kretsene, i hver bærbar PC vi undersøker.
 Men hva Gagler et al. 2022 fant var et makroskala-mønster, "som ikke direkte korrelerer med en høy grad av mikroskala-universalitet," og "kan ikke forklares direkte av universaliteten til de underliggende komponentfunksjonene." I en medfølgende nyhetsartikkel beskrev prosjektmedforfatter Chris Kempes, fra Santa Fe Institute, hovedfunnet deres i form av funksjonelle synonymer: makroskala-funksjoner er påkrevd, men ikke de identiske komponentene på lavere nivå:
Men hva Gagler et al. 2022 fant var et makroskala-mønster, "som ikke direkte korrelerer med en høy grad av mikroskala-universalitet," og "kan ikke forklares direkte av universaliteten til de underliggende komponentfunksjonene." I en medfølgende nyhetsartikkel beskrev prosjektmedforfatter Chris Kempes, fra Santa Fe Institute, hovedfunnet deres i form av funksjonelle synonymer: makroskala-funksjoner er påkrevd, men ikke de identiske komponentene på lavere nivå:
"Her finner vi at du får disse skaleringsforholdene uten å måtte bevare eksakt medlemskap. Du trenger et visst antall transferaser, men ikke spesielle transferaser, sier SFI-professor Chris Kempes, medforfatter på avisen. "Det er mange "synonymer", og disse synonymene skaleres på systematiske måter."
Som Gagler et al. ram inn punktet i selve arikkelen (uthevelse lagt til):
-Et kritisk spørsmål er om universalitetsklassene som er identifisert her, er et produkt av livets felles opphav. En begrensning ved det tradisjonelle synet på biokjemisk universalitet er at universalitet bare kan forklares i form av evolusjonær beredskap og delt historie, noe som utfordrer vår evne til å generalisere utover livets enestående opphav slik vi kjenner det. I stedet viste vi her at universalitetsklasser ikke er direkte korrelert med komponentuniversalitet, noe som indikerer at det dukker opp som en makroskopisk regularitet i storskala-statistikken over katalytisk funksjonelt mangfold. Videre kan EC-universalitet ikke bare forklares på grunn av fylogenetisk slektskap siden spekteret av totale enzymfunksjoner spenner over to størrelsesordener, noe som vitner om en bred dekning av genomisk mangfold.
Bilde 5. Ikke grunnlag for felles avstamning
Høres ut som intelligent design
 Det er interessant å merke seg at denne artikkelen ble redigert (for PNAS) av Eugene Koonin fra National Center for Biotechnology Information. I mange år har Koonin hevdet i sitt eget arbeid at den antatte "universaliteten på grunn av aner"-premisset for neo-darwinistisk teori ikke lenger holder, i stor grad på grunn av det han og andre har kalt "ikke-ortolog genforskyvning" . Dette er et gjennomgripende mønster for bruk av funksjonelle synonymer - enzymfunksjoner som utføres av forskjellige molekylære aktører - i forskjellige arter (oversetters utheving) . I 2016 skrev Koonin:
Det er interessant å merke seg at denne artikkelen ble redigert (for PNAS) av Eugene Koonin fra National Center for Biotechnology Information. I mange år har Koonin hevdet i sitt eget arbeid at den antatte "universaliteten på grunn av aner"-premisset for neo-darwinistisk teori ikke lenger holder, i stor grad på grunn av det han og andre har kalt "ikke-ortolog genforskyvning" . Dette er et gjennomgripende mønster for bruk av funksjonelle synonymer - enzymfunksjoner som utføres av forskjellige molekylære aktører - i forskjellige arter (oversetters utheving) . I 2016 skrev Koonin:
-Etter hvert som genomdatabasen vokser, blir det klart at NOGD når over de fleste funksjonelle systemer og veier slik at det er svært få funksjoner som virkelig er "monomorfe", dvs. representert av gener fra samme ortologe avstamning i alle organismer som er utstyrt med disse funksjonene. Følgelig har den universelle kjernen i livet krympet nesten til den forsvinner ... det er ingen universell genetisk kjerne i livet, på grunn av det (nær) allestedsnærværende ved "ikke-ortolog genforskyvning" .
Universelle funksjonskrav, men uten identiteten til materialkomponenter - høres ut som design.
Bilde 6. Sannsynlighet for tilfeldig dannelse: Forsvinnende liten
Oversettelse og bilder ved Asbjørn E. Lund